
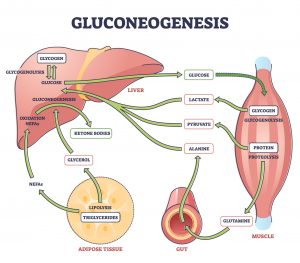
Gluconeogenesis is a vital metabolic pathway that allows organisms to maintain blood glucose levels during periods of fasting, starvation, or intense exercise. Derived from the Greek words “gluco” (sugar), “neo” (new), and “genesis” (creation), gluconeogenesis literally means “the creation of new glucose.” Unlike glycolysis, which breaks down glucose to produce energy, gluconeogenesis synthesizes glucose from non-carbohydrate precursors such as lactate, glycerol, and certain amino acids. This pathway is particularly critical for tissues that rely heavily on glucose, such as the brain, red blood cells, and renal medulla.
Gluconeogenesis is not simply a reversal of glycolysis; although it shares several enzymatic steps with glycolysis, it employs unique bypass reactions to overcome thermodynamically irreversible steps. Understanding gluconeogenesis requires a detailed appreciation of its enzymatic mechanisms, regulation, and physiological role.
1. Importance of Gluconeogenesis
The body’s dependence on glucose varies between tissues:
-
Brain: Under normal conditions, the brain consumes about 120 grams of glucose per day. During prolonged fasting, the brain adapts partially to ketone bodies but still requires some glucose.
-
Red blood cells: These cells rely exclusively on glucose for ATP production via anaerobic glycolysis.
-
Renal medulla and lens of the eye: Also dependent on glucose due to limited mitochondrial activity.
Gluconeogenesis ensures that plasma glucose levels remain within the physiological range (70–110 mg/dL) to supply these tissues. Without this pathway, hypoglycemia would quickly impair brain function and endanger life.
2. Sites of Gluconeogenesis
Gluconeogenesis primarily occurs in the liver (about 90% of systemic glucose output) and, to a lesser extent, the kidneys (especially during prolonged fasting). Skeletal muscle and adipose tissue generally do not perform gluconeogenesis because they lack the enzyme glucose-6-phosphatase, which is essential for converting glucose-6-phosphate into free glucose that can enter the bloodstream.
3. Precursors for Gluconeogenesis
Gluconeogenesis utilizes non-carbohydrate molecules as starting substrates:
-
Lactate: Produced by anaerobic glycolysis in muscle and red blood cells. Lactate is transported to the liver via the Cori cycle, where it is converted to pyruvate by lactate dehydrogenase.
-
Glycerol: Derived from the breakdown of triglycerides in adipose tissue. Glycerol is phosphorylated to glycerol-3-phosphate by glycerol kinase and then converted to dihydroxyacetone phosphate (DHAP), entering the gluconeogenic pathway.
-
Glucogenic amino acids: Amino acids such as alanine, glutamine, and others can be converted into pyruvate or intermediates of the TCA cycle (oxaloacetate, fumarate, succinyl-CoA) and used in glucose synthesis.
-
Propionate: In ruminants, propionate from gut microbial fermentation provides an additional gluconeogenic substrate.
4. Gluconeogenic Pathway
Gluconeogenesis shares many steps with glycolysis but bypasses the three thermodynamically irreversible steps of glycolysis:
-
Hexokinase/Glucokinase Step: Glucose → Glucose-6-phosphate
-
Phosphofructokinase-1 Step: Fructose-6-phosphate → Fructose-1,6-bisphosphate
-
Pyruvate Kinase Step: Phosphoenolpyruvate → Pyruvate
To bypass these steps, gluconeogenesis uses specific enzymes and reactions:
4.1 Pyruvate to Phosphoenolpyruvate (PEP)
This is a two-step process:
-
Pyruvate Carboxylase: Pyruvate is carboxylated to oxaloacetate in the mitochondria, using biotin as a cofactor and ATP as an energy source:
Pyruvate + CO2+ATP→Oxaloacetate + ADP + Pi
-
PEP Carboxykinase (PEPCK): Oxaloacetate is decarboxylated and phosphorylated using GTP to form phosphoenolpyruvate:
Oxaloacetate + GTP→PEP + GDP + CO2
This step circumvents the irreversible pyruvate kinase reaction in glycolysis.
4.2 Fructose-1,6-bisphosphate to Fructose-6-phosphate
The glycolytic enzyme phosphofructokinase-1 (PFK-1) is bypassed by fructose-1,6-bisphosphatase (FBPase-1):
Fructose-1,6-bisphosphate + H2O→Fructose-6-phosphate + Pi\text{Fructose-1,6-bisphosphate + H}_2\text{O} \rightarrow \text{Fructose-6-phosphate + Pi}
This reaction is highly regulated and essentially irreversible under cellular conditions.
4.3 Glucose-6-phosphate to Glucose
Finally, glucose-6-phosphate is converted to free glucose by glucose-6-phosphatase in the endoplasmic reticulum:
Glucose-6-phosphate + H2O→Glucose + Pi
This step allows glucose to exit the liver and kidney cells to maintain blood glucose levels.
5. Energy Requirements
Gluconeogenesis is energetically expensive. Synthesizing one molecule of glucose from pyruvate consumes:
-
4 ATP
-
2 GTP
-
2 NADH
For example, converting two molecules of pyruvate to one molecule of glucose involves:
-
Pyruvate carboxylase: 2 ATP
-
PEP carboxykinase: 2 GTP
-
Phosphoglycerate kinase: 2 ATP (for 3-phosphoglycerate → 1,3-bisphosphoglycerate)
-
Glyceraldehyde-3-phosphate dehydrogenase: 2 NADH (for reduction of 1,3-bisphosphoglycerate)
This high energy requirement explains why gluconeogenesis is tightly regulated.
6. Regulation of Gluconeogenesis
Gluconeogenesis and glycolysis are reciprocally regulated to prevent futile cycling. Regulation occurs at three major levels:
6.1 Allosteric Regulation
-
Pyruvate carboxylase: Activated by acetyl-CoA (signals high energy state and need for glucose production).
-
PEPCK: Regulated by substrate availability and hormones; generally induced by glucagon.
-
Fructose-1,6-bisphosphatase: Inhibited by AMP (signals low energy) and fructose-2,6-bisphosphate (a potent inhibitor of gluconeogenesis).
-
Glucose-6-phosphatase: Less tightly regulated but sensitive to substrate availability.
6.2 Hormonal Regulation
-
Glucagon: Released during fasting, it stimulates gluconeogenesis by:
-
Increasing cAMP → activating protein kinase A
-
Phosphorylating and activating key enzymes (e.g., PEPCK)
-
Reducing fructose-2,6-bisphosphate levels, disinhibiting FBPase-1
-
-
Insulin: Released after meals, it inhibits gluconeogenesis by decreasing PEPCK expression and increasing glycolytic enzyme activity.
-
Cortisol: A glucocorticoid hormone that enhances gluconeogenic enzyme transcription during stress or fasting.
6.3 Substrate Availability
The rate of gluconeogenesis is influenced by the levels of lactate, glycerol, and glucogenic amino acids. Increased muscle proteolysis during fasting provides alanine, which is a major substrate for hepatic gluconeogenesis.
7. Interconnection with Other Metabolic Pathways
Gluconeogenesis is integrated with several other pathways:
-
Cori Cycle: Lactate from muscle is transported to the liver, converted to pyruvate, and used for glucose synthesis.
-
Alanine Cycle: Muscle proteolysis generates alanine, which is transported to the liver, converted to pyruvate, and used in gluconeogenesis.
-
Glycerol Pathway: Triglyceride breakdown in adipose tissue provides glycerol, which enters gluconeogenesis as DHAP.
These cycles illustrate the systemic coordination of energy metabolism between tissues.
8. Physiological Significance
Gluconeogenesis is crucial for:
-
Maintaining Blood Glucose Levels: Preventing hypoglycemia during fasting, starvation, or prolonged exercise.
-
Brain Function: Providing glucose for neurons and maintaining cognitive function.
-
Red Blood Cell Function: Sustaining anaerobic glycolysis in erythrocytes.
-
Metabolic Flexibility: Allowing adaptation to changing dietary and energy conditions.
During prolonged fasting, gluconeogenesis becomes the primary source of glucose, while glycogen stores are depleted within 24 hours.
9. Clinical Relevance
Dysregulation of gluconeogenesis has significant clinical implications:
-
Diabetes Mellitus: In type 2 diabetes, excessive gluconeogenesis contributes to hyperglycemia despite high blood glucose levels. Metformin, a common antidiabetic drug, inhibits hepatic gluconeogenesis to improve glycemic control.
-
Hypoglycemia: Impaired gluconeogenesis due to liver disease, enzyme deficiencies, or starvation can lead to dangerously low blood glucose.
-
Inborn Errors of Metabolism: Deficiencies in enzymes like pyruvate carboxylase or glucose-6-phosphatase result in glycogen storage diseases and severe hypoglycemia.
10. Evolutionary Perspective
Gluconeogenesis is conserved across vertebrates because glucose is essential for vital tissues. In ruminants, propionate from gut fermentation provides a major substrate. In humans, the evolution of gluconeogenesis allows survival during periods of food scarcity, linking protein, fat, and carbohydrate metabolism efficiently.
Gluconeogenesis is an essential metabolic pathway that allows organisms to synthesize glucose from non-carbohydrate precursors. It is tightly regulated at the enzymatic, hormonal, and substrate levels to balance energy needs and prevent futile cycles. By maintaining blood glucose during fasting and starvation, gluconeogenesis supports critical physiological functions, particularly in the brain and red blood cells. Dysregulation of this pathway has significant clinical implications, highlighting its importance in health and disease.
The pathway illustrates the remarkable adaptability of human metabolism, integrating multiple tissues and metabolic substrates to maintain homeostasis. Its energetic cost reflects the high priority of glucose supply, emphasizing the delicate balance between energy expenditure and survival.
Leave a Reply